Hipótese heterotrófica sobre a origem da vida: o que é, o que diz
Entenda o que é a hipótese heterotrófica e como os primeiros seres vivos teriam surgido na Terra
Há várias explicações de como a vida teria surgido em nosso planeta e uma dessas hipóteses é da hipótese heterotrófica.
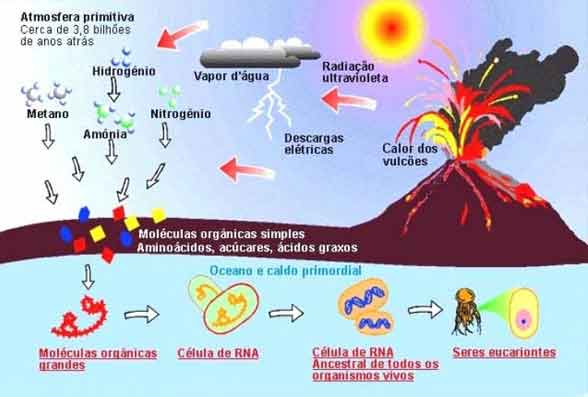
A vida na Terra começou há cerca de 3,5 bilhões de anos. Nesse ponto, no desenvolvimento da Terra, a atmosfera era muito diferente do que é hoje.
Ao contrário da atmosfera atual, que é composta principalmente nitrogênio e oxigênio, a atmosfera inicial da Terra continha principalmente hidrogênio, água, amônia e metano. Veja o vídeo logo abaixo e depois continue com a discussão.
O que é hipótese heterotrófica
A hipótese heterotrófica afirma que os primeiros seres vivos a surgirem na Terra eram organismos heterótrofos, ou seja, não produziam o próprio alimento. Os organismos tinham processos fisiológicos muito simples e provavelmente eram fermentadores.
Em experimentos, os cientistas mostraram que as descargas elétricas de raios, radioatividade e luz ultravioleta fizeram com que os elementos na atmosfera da Terra inicial formassem as moléculas básicas de química biológica, como nucleotídeos, proteínas simples e ATP. Parece provável, então, que a Terra estava coberta por uma sopa quente e fina de água e materiais orgânicos.
Ao longo do tempo, as moléculas tornaram-se mais complexas e começaram a colaborar para executar processos metabólicos. Eventualmente, as primeiras células surgiram.
Essas células eram heterótrofas, que não podiam produzir seus próprios alimentos e, em vez disso, alimentadas com o material orgânico da sopa primordial.
Daí vem o nome dessa teoria: a hipótese heterotrófica.
Os processos metabólicos anaeróbicos dos heterótrofos liberaram dióxido de carbono na atmosfera, o que permitiu a evolução dos autótrofos fotossintéticos, que poderiam usar luz e CO₂ para produzir seus próprios alimentos.
Os seres autotróficos liberaram oxigênio na atmosfera. Para a maioria dos heterótrofos anaeróbicos originais, o oxigênio mostrou-se venenoso.
Os poucos heterótrofos que sobreviveram à mudança de ambiente geralmente evoluíram para a capacidade de realizar a respiração aeróbica.
Durante os últimos bilhões de anos, os autótrofos aeróbicos e os heterótrofos tornaram-se as formas de vida dominantes no planeta e evoluíram para toda a diversidade da vida agora visível na Terra.
Segundo a hipótese heterotrófica os primeiros seres vivos seriam organismos fermentadores muito simples.
Evidência de Evolução
A humanidade sempre se perguntou sobre suas origens e as origens da vida em torno dela. Muitas culturas têm mitos de criação antiga que explicam a origem da Terra e sua vida. Nas culturas ocidentais, as ideias sobre evolução foram originalmente baseadas na Bíblia.
O livro de Gênesis relaciona como Deus criou toda a vida na Terra há cerca de 6.000 anos atrás em um evento de criação em massa.
Os defensores do criacionismo apoiam a conta Gênesis e afirmam que as espécies foram criadas exatamente como são atualmente encontradas na natureza. Esta concepção formal mais antiga da origem da vida ainda tem defensores hoje.
No entanto, há cerca de 200 anos, evidências científicas começaram a lançar dúvidas sobre o criacionismo. Esta evidência vem em uma variedade de formas.
Formação de fóssil em uma rocha
Os fósseis fornecem a única prova direta da história da evolução. A formação de fóssil ocorre quando o sedimento cobre algum material ou enche uma impressão.
Muito gradualmente, calor e pressão endurecem o sedimento e os minerais circundantes o substituem, criando fósseis. Os fósseis da vida pré-histórica podem ser ossos, conchas ou dentes que estão enterrados na rocha, e também podem ser vestígios de folhas ou pegadas deixadas por organismos.
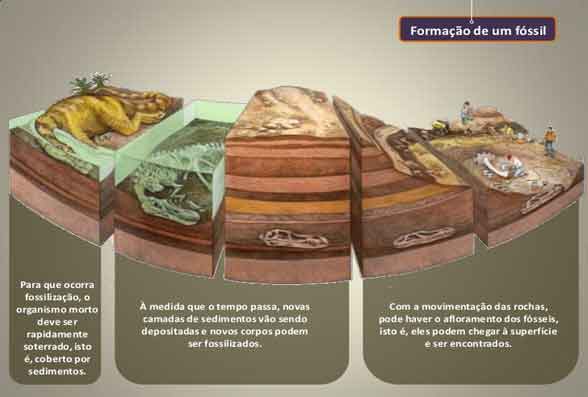
Juntos, os fósseis podem ser usados para construir um que ofereça uma linha de tempo de fósseis que reúnem a história. Para reunir o registro fóssil, os cientistas devem poder namorar os fósseis até um certo período de tempo.
Os estratos da rocha em que os fósseis são encontrados fornecem pistas sobre suas idades relativas. Se dois fósseis forem encontrados na mesma localização geográfica, mas um é encontrado em uma camada de sedimento que está abaixo da outra camada, é provável que o fóssil na camada inferior seja de uma era anterior.
Afinal, a primeira camada de sedimento já devia estar no chão para que a segunda camada começasse a se acumular. Além das camadas de sedimentos, novas técnicas, como o decaimento radioativo ou a datação por carbono também podem ajudar a determinar a idade de um fóssil.
Existem, no entanto, limitações à informação que os fósseis podem fornecer. Antes de tudo, a fossilização é um evento improvável.
Na maioria das vezes, os restos e outros vestígios de organismos são esmagados ou consumidos antes de serem fossilizados. Além disso, os fósseis só podem formar-se em áreas com rochas sedimentares, como os pisos oceânicos.
Os organismos que vivem nesses ambientes são, portanto, mais propensos a se tornar fósseis. Finalmente, a erosão de superfícies expostas ou movimentos geológicos, como terremotos, pode destruir fósseis já formados. Todas essas condições levam a grandes e numerosas lacunas no registro fóssil.
Anatomia Comparada
Os cientistas muitas vezes tentam determinar a relação de dois organismos através da comparação de estruturas externas e internas.
O estudo da anatomia comparada é uma extensão do raciocínio lógico de que organismos com estruturas similares devem ter adquirido esses traços de um antepassado comum. Por exemplo, o flipper de uma baleia e um braço humano parecem ser bastante diferentes quando olhamos de fora.
Mas a estrutura óssea de cada um é surpreendentemente similar, sugerindo que as baleias e os humanos têm um caminho ancestral comum na pré-história.
Características anatômicas em diferentes espécies que apontam para um antepassado comum são chamadas de estruturas homólogas .
No entanto, anatomistas comparativos não podem simplesmente assumir que toda estrutura similar aponta para uma origem evolutiva comum.
Um anatomista comparativo precipitado e imprudente pode assumir que os morcegos e os insetos compartilham um antepassado comum, uma vez que ambos têm asas.
Mas um olhar mais atento sobre a estrutura das asas mostra que há muito pouco em comum entre eles além de sua função.
Na verdade, a ala de morcego está muito mais próxima em estrutura para o braço de um homem e a barbatana de uma baleia do que para as asas de um inseto.
Em outras palavras, os morcegos e os insetos evoluíram sua capacidade de voar ao longo de dois caminhos evolutivos muito separados.
Esses tipos de estruturas, que têm semelhanças superficiais por causa da semelhança de função, mas não resultam de um antepassado comum, são chamadas de estruturas análogas.
Além das estruturas homólogas e análogas, as estruturas vestigiais, que não servem de função moderna aparente, podem ajudar a determinar como um organismo pode ter evoluído ao longo do tempo.
Em humanos, o apêndice é inútil, mas em vacas e outros herbívoros de mamíferos, uma estrutura similar é usada para digerir a celulose.
A existência do apêndice sugere que os humanos compartilham uma ascendência evolutiva comum com outros herbívoros de mamíferos.
O fato de que o apêndice agora não serve para nada em seres humanos demonstra que os humanos e os herbívoros de mamíferos há muito tempo divergiram em seus caminhos evolutivos.
Embriologia Comparada
Estruturas homólogas não presentes em organismos adultos muitas vezes não aparecem de alguma forma durante o desenvolvimento embrionário.
As espécies que têm pouca semelhança entre si em suas formas adultas podem ter estágios embrionários surpreendentemente semelhantes.
De certa forma, é quase como se o embrião passasse por muitos estágios evolutivos para produzir o organismo maduro. Por exemplo, para uma grande parte do seu desenvolvimento, o embrião humano possui uma cauda, bem como aqueles de nossos parentes próximos de primatas.
Esta cauda geralmente é reabsorvida antes do nascimento, mas ocasionalmente nascem crianças com a estrutura ancestral intacta.
Embora não estejam geralmente presentes no organismo adulto, as caudas podem ser consideradas características homólogas entre humanos e primatas.
Em geral, as espécies mais estreitamente relacionadas são, mais seus processos embriológicos de desenvolvimento se assemelham uns aos outros.
Evolução Molecular
Assim como a anatomia comparativa é usada para determinar a relação anatômica das espécies, a biologia molecular pode ser usada para determinar as relações evolutivas no nível molecular.
Duas espécies que estão intimamente relacionadas terão menos diferenças genéticas ou proteicas entre elas do que duas espécies que estão distantes e divididas no desenvolvimento evolutivo há muito tempo no passado.
Certos genes ou proteínas em organismos mudam a uma taxa constante ao longo do tempo. Esses genes e proteínas, chamados relógios moleculares, porque são tão constantes em sua taxa de mudança, são especialmente úteis na comparação da evolução molecular de diferentes espécies.
Os cientistas podem usar a taxa de mudança no gene ou proteína para calcular o ponto em que duas espécies finalmente compartilharam um antepassado comum.
Por exemplo, o RNA ribossômico tem uma taxa de mudança muito lenta, por isso é comumente usado como um relógio molecular para determinar relacionamentos entre espécies extremamente antigas.
O citocromo c, uma proteína que desempenha um papel importante na respiração aeróbia, é um exemplo de uma proteína comumente usada como um relógio molecular.
Teorias da evolução
No século XIX, como evidências crescentes sugeriram que as espécies mudaram ao longo do tempo, os cientistas começaram a desenvolver teorias para explicar como essas mudanças surgem. Durante este tempo, houve duas teorias notáveis da evolução. O primeiro, proposto por Lamarck, acabou por ser incorreto. O segundo, desenvolvido por Darwin, é a base de toda a teoria evolutiva.
Lamarck: uso e desuso
A primeira teoria notável da evolução foi proposta por Jean-Baptiste Lamarck (1744-1829). Ele descreveu um mecanismo de duas partes pelo qual a mudança evolutiva foi gradualmente introduzida nas espécies e transmitida através de gerações. Sua teoria é referida como a teoria da transformação ou Lamarckismo.
O exemplo clássico usado para explicar o Lamarckismo é o pescoço alongado da girafa. De acordo com a teoria de Lamarck, uma dada girafa poderia, ao longo de uma vida de esforço para alcançar ramos altos, desenvolver um pescoço alongado. Isso ilustra vividamente a crença de Lamarck de que o uso poderia amplificar ou aprimorar uma característica.
Da mesma forma, ele acreditava que o desuso causaria uma redução da característica. De acordo com a teoria de Lamarck, as alas de pinguins, por exemplo, eram compreensivelmente menores do que as asas de outras aves porque os pinguins não usavam suas asas para voar.
A segunda parte do mecanismo de evolução de Lamarck envolveu a herança de caracteres adquiridos. Ele acreditava que se os traços de um organismo mudassem ao longo de sua vida, o organismo passaria esses traços para a sua prole.
A teoria de Lamarck foi comprovada erroneamente em suas duas premissas básicas. Primeiro, um organismo não pode mudar sua estrutura de forma fundamental através do uso ou do desuso.
O pescoço de uma girafa não se tornará mais longo ou mais curto, alongando as folhas. Em segundo lugar, a genética moderna mostra que é impossível transmitir os traços adquiridos; os traços que um organismo pode transmitir são determinados pelo genótipo de suas células sexuais, o que não muda de acordo com as mudanças no fenótipo.
Darwin: seleção natural
Enquanto navegava a bordo do HMS Beagle , o inglês Charles Darwin teve a oportunidade de estudar a vida selvagem das Ilhas Galápagos. Nas ilhas, ele ficou maravilhado com a grande diversidade da vida.
Mais particularmente, ele se interessou pelos vários tentilhões das ilhas, cujos bicos foram altamente adaptados aos seus estilos de vida particulares.
Ele colocou a hipótese de que deve haver algum processo que criou essa diversidade e adaptação, e ele passou a maior parte do tempo tentando descobrir exatamente qual seria o processo.
Em 1859, ele publicou sua teoria da seleção natural e a evolução que produziu. Darwin explicou sua teoria através de quatro pontos básicos:
- Cada espécie produz mais prole do que pode sobreviver.
- Os organismos individuais que compõem uma população maior nascem com certas variações.
- A superabundância da prole cria uma competição para a sobrevivência entre organismos individuais. Os indivíduos que apresentam as variações mais favoráveis sobreviverão e se reproduzirão, enquanto aqueles com variações menos favoráveis são menos propensos a sobreviver e reproduzir.
- As variações são passadas do pai para a prole.
A seleção natural cria mudanças dentro de uma espécie através da competição, ou a luta pela vida. Membros de uma espécie competem uns com os outros e com outras espécies para recursos.
Nesta competição, os indivíduos que são mais adequados – os indivíduos que têm certas variações que os tornam mais adaptados aos seus ambientes – são os mais capazes de sobreviver, reproduzir e passar seus traços para a sua prole. A competição que a teoria de Darwin descreve às vezes é chamada de sobrevivência do mais apto.
Seleção natural em ação
Um dos melhores exemplos de seleção natural é uma história verdadeira que aconteceu na Inglaterra em torno da virada do século. Perto de uma cidade agrícola, viveu uma espécie de traça. A traça passou grande parte de seu tempo empoleirada no casco coberto de líquen das árvores da área.
A maioria das mariposas era de cor de pimenta, embora algumas delas fossem pretas. Quando as traças de cor de pimenta estavam ligadas à casca coberta de líquen das árvores na região, era bastante difícil para os predadores vê-los. As mariposas pretas eram fáceis de detectar contra os troncos salpicados de preto e branco.
A cidade vizinha, no entanto, lentamente se industrializou. Fumaças e fundições da cidade incharam fuligem e fumam no ar. Em pouco tempo, o fulgor estabeleceu tudo, incluindo as árvores, e matou muito do líquen.
Como resultado, a aparência das árvores tornou-se quase de cor preta. De repente, as mariposas de cor de pimenta eram óbvias contra os troncos das árvores escuras, enquanto as mariposas pretas que tinham sido fáceis de detectar agora se misturavam contra as árvores.
Ao longo dos anos, os moradores da cidade notaram que a população das mariposas mudou. Enquanto cerca de 90 por cento das mariposas costumavam ser leves, depois que as árvores ficaram pretas, a população da traça ficou cada vez mais preta.
Quando as árvores eram de cor mais clara, a seleção natural favoreceu as mariposas de cor de pimenta, porque essas mariposas eram mais difíceis de detectar. Como resultado, as mariposas de cor de pimenta viviam para se reproduzir e tinham descendentes de cor de pimenta, enquanto muito menos das mariposas pretas viviam para produzir descendentes pretas.
Quando a indústria na cidade matou o líquen e cobriu as árvores em fuligem, no entanto, a pressão de seleção mudou. De repente, as traças pretas eram mais propensas a sobreviver e a ter descendência.
Em cada geração, mais mariposas negras sobreviveram e tiveram descendência, enquanto poucas mariposas mais claras sobreviveram para ter descendentes. Ao longo do tempo, a população como um todo evoluiu da maior parte da cor branca até a maioria na cor preta.
Tipos de Seleção Natural
Em uma população normal sem pressão de seleção, traços individuais, como altura, variam na população. A maioria dos indivíduos tem uma altura média, enquanto menos são extremamente baixas ou extremamente altas. A distribuição de altura cai em uma curva de sino.
A seleção natural pode operar nesta população de três formas básicas.
Seleção de estabilização
A seleção de estabilização elimina indivíduos extremos. Uma planta que é muito curta pode não ser capaz de competir com outras plantas para a luz solar.
No entanto, plantas extremamente altas podem ser mais suscetíveis a danos causados pelo vento. Combinadas, estas duas pressões de seleção agem para favorecer plantas de estatura média.
Seleção direcional
Seleção direcional seleciona contra um extremo. No exemplo familiar de pescoço de girafas, houve uma pressão de seleção contra pescoços curtos, uma vez que indivíduos com pescoço curto não conseguiam alcançar tantas folhas para se alimentar. Como resultado, a distribuição do comprimento do pescoço mudou para favorecer pessoas com pescoço longo.
Seleção disruptiva
A seleção disruptiva elimina indivíduos intermediários. Por exemplo, imagine uma planta de altura extremamente variável que seja polinizada por três insetos polinizadores diferentes: um que foi atraído por plantas curtas, outra que preferia plantas de estatura média e uma terceira que visitava apenas as plantas mais altas.
Se o polinizador que preferisse plantas de altura média desapareceram de uma área, as plantas de altura média seriam selecionadas contra, e a população tenderia para plantas altas e altas, mas não plantas de estatura média.
A base genética para a evolução
A teoria de Darwin sobre a seleção natural e evolução baseia-se em duas idéias cruciais:
- Existem variações nos indivíduos dentro de uma população.
- Essas variações são passadas de uma geração para a próxima.
Mas Darwin não tinha ideia de como essas variações eram ou como elas passaram de uma geração para outra. As experiências de Mendel e o desenvolvimento da ciência da genética forneceram respostas.
A genética explica que o fenótipo – os atributos físicos de um organismo – é produzido pelo genótipo de um organismo.
Através do mecanismo das mutações, a genética explica como as variações surgiram entre os indivíduos na forma de diferentes genes alelos. A meiose, a reprodução sexual e a herança dos alelos explicam como as variações entre os organismos são transmitidas do pai para a prole.
Com a moderna compreensão de genes e heranças, é possível redefinir a seleção natural e a evolução em termos genéticos. Os alelos particulares que um organismo herda de seus pais determinam os atributos físicos do organismo e, portanto, sua aptidão para a sobrevivência.
Quando as forças da seleção natural resultam na sobrevivência do mais apto, o que essas forças realmente estão fazendo é selecionar quais alelos serão transmitidos de uma geração para outra.
Uma vez que você vê essa seleção natural é realmente uma seleção da passagem de alelos de geração para geração, você pode ver ainda que as forças da seleção natural podem mudar a frequência de cada alelo particular dentro do pool de genes de uma população, que é a soma total de todos os alelos dentro de uma determinada população.
Usando a genética, pode-se criar uma nova definição de evolução como a mudança nas frequências de alelos no grupo gene de uma população ao longo do tempo.
Por exemplo, na população de mariposas que discutimos anteriormente, depois que as árvores escureceram, a freqüência dos alelos para coloração negra aumentou no pool de genes, enquanto a freqüência de alelos para coloração leve diminuiu.
Equilíbrio Hardy-Weinberg
O princípio de Hardy-Weinberg afirma que uma população de reprodução sexual terá frequências alélicas estáveis e, portanto, não passará por evolução, tendo em conta as seguintes cinco condições:
- grande tamanho da população
- sem imigração ou emigração
- acasalamento aleatório
- sucesso reprodutivo aleatório
- sem mutação
O princípio Hardy-Weinberg prova que a variabilidade e a herança por si só não são suficientes para causar a evolução; A seleção natural deve impulsionar a evolução.
Uma população que atende a todas essas condições é dito estar no equilíbrio de Hardy-Weinberg . Poucas populações naturais já experimentaram o equilíbrio de Hardy-Weinberg, porém, uma vez que as populações grandes raramente são encontradas isoladamente, todas as populações experimentam algum nível de mutação e a seleção natural simplesmente não pode ser evitada.
Desenvolvimento de novas espécies (especiação)
A definição científica de uma espécie é um grupo discreto de organismos que só podem se reproduzir dentro dos seus próprios limites. Em outras palavras, os membros de uma espécie não podem se cruzar com os membros de outra espécie.
Espera-se que cada espécie experimente isolamento reprodutivo. Se você pensa sobre a evolução em termos de genética, essa definição de espécie faz muito sentido: se as espécies pudessem se cruzar, elas poderiam compartilhar o fluxo de genes e sua evolução não seria separada. Mas, como as espécies não podem se cruzar, cada espécie existe em seu próprio caminho individual.
À medida que as populações mudam, novas espécies evoluem. Esse processo é conhecido como especiação. Através da especiação, os primeiros organismos simples foram capazes de se ramificar e povoar o mundo com milhões de espécies diferentes.
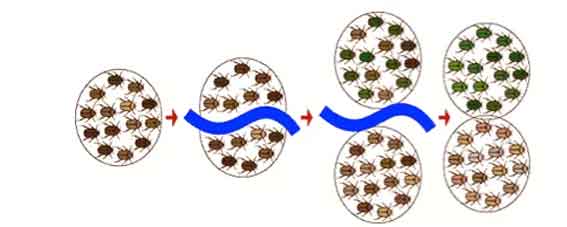
A especificação também é chamada de evolução divergente, uma vez que uma nova espécie se desenvolve, diverge de uma forma anterior. Todos os traços homólogos são produzidos por evolução divergente.
Baleias e humanos compartilham um antepassado comum distante. Através da especiação, esse antepassado sofreu uma evolução divergente e deu origem a novas espécies, o que, por sua vez, deu origem a novas espécies, que ao longo de milhões de anos resultaram em baleias e humanos.
O antepassado original tinha uma estrutura de membros que, ao longo de milhões de anos e ocorrências sucessivas de evolução divergente, evoluiu para a barbatana da baleia e para o braço do humano.
A especiação ocorre quando duas populações se tornam isoladas reprodutivamente. Uma vez que o isolamento reprodutivo ocorre para uma nova espécie, ele começará a evoluir de forma independente. Existem duas maneiras principais pelas quais a especiação pode ocorrer.
A especiação alopática ocorre quando as populações de uma espécie se tornam geograficamente isoladas para que elas não possam se cruzar.
Ao longo do tempo, as populações podem se tornar geneticamente diferentes em resposta às pressões de seleção únicas que operam em seus diferentes ambientes.
Eventualmente, as diferenças genéticas entre as duas populações tornar-se-ão tão extremas que as duas populações não poderiam se cruzar, mesmo que a barreira geográfica tenha desaparecido.
Uma segunda forma mais comum de especiação é a radiação adaptativa, que é a criação de várias novas espécies de uma única espécie-mãe. Pense em uma população de uma determinada espécie, que nomearemos imaginativamente a população 1.
A população se move para um novo habitat e se estabelece em um nicho ou papel no habitat (discutimos nichos com mais detalhes no capítulo sobre Ecologia).
Ao fazê-lo, ele se adapta ao seu novo ambiente e torna-se diferente das espécies-mãe. Se uma nova população das espécies-mãe, a população 2, se desloca para a área, ela também tentará ocupar o mesmo nicho que a população 1.
A competição entre a população 1 e a população 2 ocorre, pressionando ambos os grupos para se adaptar a nichos separados, distingui-los mais uns dos outros e as espécies-mãe.
Como isso acontece muitas vezes em um determinado habitat, várias espécies novas podem ser formadas a partir de uma única espécie mãe em um período relativamente curto.
A imensa diversidade de tentilhões que Darwin observou nas Ilhas Galápagos é um excelente exemplo dos produtos de radiação adaptativa.
Evolução convergente
Quando espécies diferentes habitam ambientes semelhantes, eles enfrentam pressões de seleção semelhantes ou usam partes de seus corpos para executar funções semelhantes.
Essas semelhanças podem fazer com que as espécies evoluam traços semelhantes, em um processo chamado evolução convergente.
De viver nas regiões frágeis, aquosas e árticas, onde a maioria dos alimentos existe subaquática, pinguins e baleias assassinas evoluíram algumas características semelhantes: ambos são simplificados para ajudá-los a nadar mais rapidamente debaixo d’água, ambos possuem camadas de gordura para mantê-los aquecidos, ambos têm coloração branca e preta similar que os ajuda a evitar a detecção, e ambos desenvolveram barbatanas (ou nadadeiras) para impulsioná-las através da água.
Todos esses traços semelhantes são exemplos de traços análogos, que são o produto da evolução convergente.
A evolução convergente soa como se fosse o oposto da evolução divergente, mas isso não é verdade. A evolução convergente é apenas superficial.
Do lado de fora, a barbatana de uma baleia pode parecer a flipper de um pinguim, mas a estrutura óssea de uma barbatana de baleia ainda é mais parecida com os membros de outros mamíferos do que com a estrutura das aletas de pinguins.
Mais importante ainda, a evolução convergente nunca resulta em duas espécies que ganham a capacidade de cruzar; A evolução convergente não pode levar duas espécies e transformá-las em uma.
Resumo da aula hipótese heterotrófica
Nessa aula nos vimos
- O que é hipótese heterotrófica
- Quais as teorias que explicam a origem da vida
- Anatomia Comparada
- Embriologia Comparada
- Teorias da evolução
- Evolução Molecular
- Tipos de Seleção Natural
- A base genética para a evolução
- Críticas a hipótese heterotrófica
Leitura Sugerida
Você sabia que tem bastante gente que envia posts, artigos, ou algo qualquer texto relacionado a ciências, educação ou biologia? Você gostaria de participar? Entre em contato.
Se você encontrar algo indevido nos textos, nas imagens, vídeos que recebemos, por favor, relate nos comentários que assim que puder será feita uma revisão.
Bibliografia
Universidade de São Paulo – Contribuição de conceitos químicos ao estudo da origem da vida na disciplina de Biologia – Acesso em 13 de janeiro de 2021