Organização celular dos seres vivos. As células são as unidades básicas da vida; pequenas máquinas que facilitam e sustentam cada processo dentro de um organismo vivo.
As células musculares se contraem para manter um batimento cardíaco e nos permitem mover-se, os neurônios formam redes que dão origem a memórias e permitem processos de pensamento.
As células epiteliais providenciam para formar barreiras superficiais entre os tecidos e as muitas cavidades em todo o corpo.
O que é uma organização celular
Organização celular diz respeito a relação das organelas e de como os seus componentes estão organizados em seu interior. No interior de cada célula acontecem as reações químicas necessárias para a manutenção e desenvolvimento da vida.
Não só os diferentes tipos de células facilitam funções únicas, mas suas composições moleculares, genéticas e estruturais também podem diferir.
Por esse motivo, diferentes tipos de células geralmente possuem variações no fenótipo, como o tamanho e a forma das células. Na imagem abaixo você pode ver diferentes tipos celulares dos seres humanos.
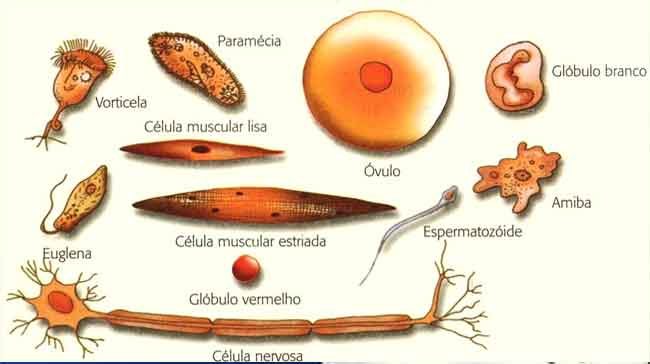
Função das células
A função de uma célula é alcançada através do ponto culminante de centenas de processos menores, muitos dos quais são dependentes uns dos outros e compartilham proteínas ou componentes moleculares.
Apesar das variações fenotípicas e funcionais que existem entre os tipos de células, é verdade que existe um alto nível de similaridade ao explorar os processos subcelulares, os componentes envolvidos e, principalmente, a organização desses componentes.
Com a maioria dos processos subcelulares sob controle regulatório preciso de outros processos subcelulares, e com componentes geralmente compartilhados entre diferentes caminhos moleculares e cascatas proteicas, a organização celular é de grande importância.
Isso é verdade para cada tipo de célula, com compartimentação de processos subcelulares, e localização de proteínas, recrutamento e entrega, garantindo que sejam constantemente repetidos de forma eficiente e com resultados precisos.
Veja também:
Como é a organização celular dos seres vivos
Ao nível básico, as células eucarióticas podem ser descritas como contendo três regiões sub-celulares distintas; nomeadamente a membrana, o citosol e o núcleo. Contudo, a compartimentação celular é ainda mais complicada pela abundância de organelas específicas.
Apesar de ter apenas vários nanômetros de largura, as membranas celulares são altamente enriquecidas em receptores de sinalização, proteínas transmembranares, bombas e canais e, dependendo da maquiagem, podem recrutar e reter um conjunto de proteínas importantes no campo da mecanobiologia.
Em muitos casos, esses proteínas interagem com o citoesqueleto , que reside na proximidade da membrana. O citosol, por outro lado, abriga organelas celulares, incluindo o complexo golgiense, o retículo endoplasmático (RE), ribossomos e numerosas vesículas e vacúolos. Podem existir proteínas solúveis nesta região.
Enquanto isso, o núcleo abriga o material genético e todos os componentes relacionados à sua expressão e regulação. Embora os processos do núcleo não estejam tão bem estabelecidos em termos de seu papel na mecanobiologia, os achados recentes indicam várias conexões importantes, muitas vezes com as vias de sinalização de mecanotransdução que culminam em alterações na expressão gênica.
Cada uma dessas regiões sub-celulares deve funcionar de forma coerente para a sobrevivência e o funcionamento eficiente da célula. A organização adequada de organelas, proteínas e outras moléculas em cada região permite que os componentes de proteínas individuais funcionem de forma harmônica, gerando efetivamente processos subcelulares individuais que culminam em uma função celular global.
Compartimentalização em células
A compartimentalização é um importante fator na organização celular.
As células não são uma mistura amorfa de proteínas, lipídios e outras moléculas. Em vez disso, todas as células são constituídas por compartimentos bem definidos, cada um especializado em uma função particular.
Em muitos casos, os processos subcelulares podem ser descritos com base na ocorrência na membrana plasmática , no citosol ou dentro de organelas ligadas à membrana, como o núcleo, o aparelho de Golgiense ou mesmo os componentes vesiculares do sistema de tráfico de membrana , como os lisossomos e os endossomas.
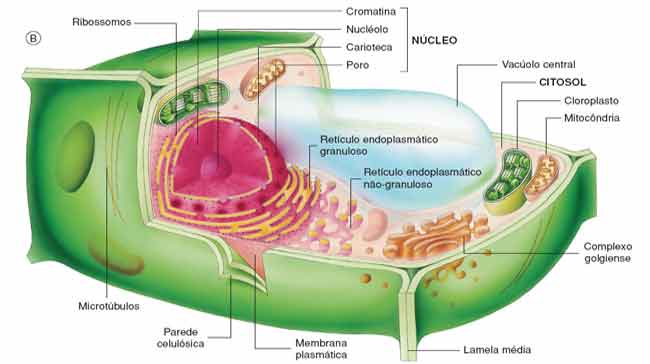
A compartimentação aumenta a eficiência de muitos processos subcelulares concentrando os componentes necessários em um espaço confinado dentro da célula.
Quando uma condição específica é necessária para facilitar um determinado processo subcelular, isso pode ser localmente contido de modo a não interromper a função de outros compartimentos subcelulares.
Por exemplo, os lisossomos requerem um pH mais baixo para facilitar a degradação do material internalizado. As bombas de protões ligadas à membrana presentes no lipossoma mantém esta condição.
Da mesma forma, uma grande área de superfície da membrana é requerida pelas mitocôndrias para gerar eficientemente ATP a partir de gradientes de elétrons em sua bicamada lipídica. Isto é conseguido através da composição estrutural deste organelo particular.
Importante, organelas individuais podem ser transportadas por toda a célula e isso localiza essencialmente todo o processo subcelular para regiões onde são necessárias.
Isso foi observado em neurônios, que possuem processos axonais extremamente longos e requerem mitocôndrias para gerar ATP em vários locais ao longo do axônio. Seria ineficiente confiar na difusão passiva do ATP ao longo do axônio.
A compartimentação também pode ter importantes implicações fisiológicas. Por exemplo, as células epiteliais polarizadas, que possuem membranas apicais e basolaterais distintas, podem, por exemplo, produzir uma superfície secretora para várias glândulas.
Da mesma forma, as células neuronais desenvolvem redes efetivas devido à produção de dendritos e processos axonais a partir de extremidades opostas do corpo celular.
Além disso, no caso de células estaminais embrionárias, a polarização celular pode resultar em destinos distintos das células filhas.
Com cada organela facilitando sua própria função, eles podem ser considerados compartimentos subcelulares por direito próprio. No entanto, sem um fornecimento regular de componentes para o compartimento, os processos e mecanismos que produzem sua função geral serão impedidos.
Com muitas proteínas e componentes moleculares que participam em múltiplos processos subcelulares e, portanto, exigidos em vários compartimentos subcelulares, o transporte efetivo da proteína e dos componentes moleculares, seja por difusão passiva ou recrutamento direcionado, é essencial para a função geral da célula.
Em seres eucariontes, a síntese de DNA, RNA, proteínas e lipídios é realizada de forma espaciotemporal. Cada molécula é produzida dentro de organelas ou compartimentos especializados com mecanismos regulatórios rígidos existentes para controlar o tempo e a taxa de síntese.
Esses mecanismos regulatórios são complicados e podem envolver loops de feedback, estímulos externos e uma multiplicidade de caminhos de sinalização.
DNA e RNA são ambos produzidos dentro do núcleo e são chamados de ácidos nucleicos.
O DNA é inteiramente replicado durante a fase s do ciclo celular. Uma cópia é então passada para cada uma das células filhas.
Durante outras fases do ciclo celular, uma quantidade mínima de DNA é sintetizada, principalmente para o reparo do material genético.
Embora uma taxa basal de síntese de RNA mantenha a síntese de mRNA ao longo da vida da célula, o mRNA para genes específicos só pode ser expresso ou pode ser regulado, após a detecção de certos sinais mecânicos ou químicos.
Como resultado, diferentes células têm diferentes perfis de mRNA, e isso geralmente é observado através do uso de tecnologias que exibem os perfis genéticos das células.
Depois de ser processado e modificado no núcleo, o mRNA transcrito é entregue ao citosol para tradução ou síntese proteica. Semelhante à síntese de RNA, um nível básico de síntese de proteína é mantido durante toda a vida da célula.
Porém, isso também pode ser alterado quando determinados estímulos induzem a produção de proteínas específicas, ou quando mecanismos regulatórios reduzem a produção de outros.
Por exemplo, a síntese de proteínas é regulada para cima durante a fase G1 do ciclo celular, imediatamente antes da fase S. Isto é para garantir que a célula tenha uma concentração suficiente da maquinaria proteica necessária para realizar a replicação do DNA e a divisão celular.
Nos procariontes, onde não há compartimentos separados, tanto a transcrição quanto a tradução ocorrem simultaneamente.
Os lipídios, que são sintetizados no retículo endoplasmático (RE) ou no complexo golgiense, são transportados para outras organelas sob a forma de vesículas que se fundem com a organela aceitadora.
Algumas células também podem usar proteínas transportadoras para transportar lipídios de um local para outro. A síntese lipídica também é dinâmica, e pode ser regulada até a proliferação celular ou durante processos que envolvem a extensão da membrana plasmática, quando novas membranas são necessárias.
Localização de Proteínas
Para que os processos celulares sejam realizados dentro de compartimentos definidos ou regiões celulares, devem existir mecanismos para garantir que os componentes proteicos necessários estejam presentes nos locais e a uma concentração adequada.
A acumulação de uma proteína em um determinado local é conhecida como localização de proteínas e é fundamental para a organização celular.
O recrutamento de proteínas é essencialmente uma forma de reconhecimento de proteínas, possibilitado pela presença de sequências específicas de aminoácidos dentro da estrutura proteica.
Por exemplo, muitas proteínas ligadas à membrana possuem péptidos de sinal que são reconhecidos pelos receptores de sinal que os orientam para o site alvo.
O sinal de localização nuclear é um desses exemplos. As proteínas que são destinadas ao retículo endoplasmático também possuem um péptido sinal.
Em outros casos, as proteínas podem transportar um remendo de sinal. Isso geralmente consiste em cerca de 30 aminoácidos que não estão presentes em uma sequência linear, mas estão em proximidade espacial próxima no espaço tridimensional.
Curiosamente, a organização de uma célula e suas várias regiões desempenham um papel na direção do recrutamento de proteínas para um determinado site.
Por exemplo, nas células epiteliais, que são polarizadas, a composição proteica na membrana apical é muito diferente daquela na membrana basolateral.
Isto é conseguido através do reconhecimento de sequências de sinais distintas que visam proteínas para cada uma dessas regiões. Por exemplo, as proteínas da membrana apical são muitas vezes ancoradas ao GPI , enquanto que as proteínas basolaterais possuem sequências de assinaturas baseadas em aminoácidos diLeu (N, N-Dimetil Leucina) ou tirosina com base em aminoácidos.
Entenda melhor do assunto acessando a aula composição química das células.
Entrega Direta de Componentes
A localização das proteínas pode resultar do reconhecimento de proteínas ou complexos solúveis de difusão passiva, no entanto, isso pode não garantir uma concentração suficiente de componentes para manter um determinado processo.
Isso pode impedir a sua conclusão, particularmente quando realizada em regiões com um volume citoplasmático limitado, como a ponta de um filopodia, ou quando os componentes são rapidamente transferidos.
Uma maneira mais eficiente de manter a concentração de componentes proteicos é por meio de sua entrega dirigida através da rede do citoesqueleto.
O citoesqueleto, composto por filamentos de actina e microtúbulos, abrange toda a célula e conecta a membrana plasmática ao núcleo e outras organelas.
Esses filamentos realizam muitos propósitos, desde o suporte estrutural até a célula, para gerar as forças necessárias para a translocação celular.
Eles também podem servir como “trilhas” nas quais as proteínas motoras podem transladar enquanto transportam carga de um local para outro; análogo a um trem de carga que transporta carga ao longo de uma rede de trilhos ferroviários.
A entrega de componentes é principalmente facilitada por motores moleculares com ATP / GTP, como miosina V ou miosina X , Cinesina ou Dineína.
Essas proteínas ou homólogos deles foram observados em uma grande quantidade de tipos celulares, incluindo leveduras, célula vegetal e célula animal.
Os motores moleculares dineína e cinesina caminham sobre os microtúbulos enquanto a miosina caminha nos filamentos de actina.
Imperativamente, esses motores caminham de maneira unidirecional, embora não necessariamente na mesma direção uns dos outros.
O transporte baseado em microtúbulos foi estudado principalmente em células neuronais. Os exons podem ter vários mícrons de comprimento (às vezes até mesmo medidores de comprimento), por isso é necessário transportar proteínas, lipídios, vesículas sinápticas, mitocôndrias e outros componentes ao longo do axônio.
Todos os microtúbulos nos axônios são unidirecionais, com extremidades “menos” que apontam para o corpo da célula e ‘mais’ que apontam para a sinapse.
Os motores de cinesina se movem ao longo dessas trilhas para transportar a carga do corpo da célula para o axônio.
A interrupção do transporte de carga mediada por cinesina está correlacionada com várias doenças neuro-musculares, como a atrofia muscular espinhal e a atrofia muscular espinhal e bulbar. A dineina, por outro lado, desempenha um papel importante no tráfico de carga em dendritos.
Caminhos de comunicação
Com diferentes processos sendo realizados em compartimentos subcelulares separados, organizados em diferentes regiões da célula, a comunicação intracelular é primordial para organização celular.
Essa comunicação, que é descrita em maior detalhe sob “sinalização celular”, permite às células manter a concentração de proteínas específicas e dentro das regiões corretas, dependendo dos requisitos de um determinado processo ou estado celular.
Isso, em última instância, garante que os compartimentos individuais funcionem de forma eficiente e permite que um processo subcelular conduza outro. Isso, em última instância, permite que uma célula facilite suas funções primárias de forma eficiente e coerente.
As vias de sinalização podem conter um sinal que se origina de fora de uma célula ou de vários compartimentos e geralmente envolve a translocação de íons, solutos, proteínas e mensageiros secundários.
Todas as células possuem receptores de superfície e outras proteínas para facilitar a detecção de sinais do ambiente extracelular.
Esses sinais podem ser na forma de íons, moléculas pequenas, péptidos, tensão de cisalhamento, forças mecânicas, calor, etc.
Uma vez que o sinal é detectado pelo receptor de superfície, ele é transmitido ao citoplasma geralmente por meio de mudança conformacional no receptor ou mudança no seu estado de fosforilação no lado citosólico.
Isso, por sua vez, desencadeia uma cascata de sinalização a jusante, que muitas vezes culmina no núcleo. O sinal geralmente resulta em mudança no perfil de expressão gênica das células, auxiliando-as a responder ao estímulo.
Você sabia que tem bastante gente que envia posts, artigos, ou algo qualquer texto relacionado a ciências, educação ou biologia? Você gostaria de participar? Entre em contato.
Se você encontrar algo indevido nos textos, nas imagens, vídeos que recebemos, por favor, relate nos comentários que assim que puder será feita uma revisão.
Referências:
CK-12 – Cell Organization – Acessado em 13 de janeiro de 2021
{kind=link}
A biologia é realemnte lnda
né isso?
respostas da unidade 1 de ciencias biologicas
muito obrigada o video me ajudou muito!!!
Tanks!!!…