O que é Metabolismo Energético
É no metabolismo energético através das reações químicas que permitem que as células produzam novas moléculas para construir sua estrutura, como a membrana citoplasmática ou o núcleo.
O metabolismo energético é o processo de produção geração de energia através da molécula de (ATP) usando para isso a energia contida nos nutrientes. O metabolismo compreende uma série de vias e processos que podem funcionar na presença de oxigênio ou na ausência.
Dois mecanismos opostos constituem o metabolismo. O primeiro é o catabolismo para extrair energia dos nutrientes. O segundo é o metabolismo celular : são efeitos anabólicos, devido a reações de síntese dos elementos constituintes essenciais à estrutura e atividade das células.
Esse processo pode ainda ser definido como um conjunto de reações químicas ao nível das células do corpo, compreende na sua totalidade três grandes vias bioquímicas para a síntese de Trifosfato de adenosina (ATP) operando simultaneamente em diferentes proporções dependendo da atividade física e/ou desportiva praticada: fosforilcreatina (via anaeróbica), a glicólise (via anaeróbica láctica) e a cadeia respiratória ao nível das mitocôndrias (via aeróbica).
O metabolismo energético no citoplasma
Ao entrar na célula, ainda no citoplasma, a molécula de glicose, que tem seis carbonos (C6H12O6), é convertida em duas moléculas de um composto com apenas três carbonos, o ácido pirúvico (C3H4O3), com sobra de elétrons e liberação de energia, que é utilizada para gerar 2 moléculas de ATP (pela união de fósforo inorgânico, Pi, ao ADP).
Esse processo, denominado glicólise não consome oxigênio e apresenta baixo rendimento energético. Além das duas moléculas de ATP, formam-se também duas moléculas de NADH, que poderão doar seus elétrons a outros compostos.
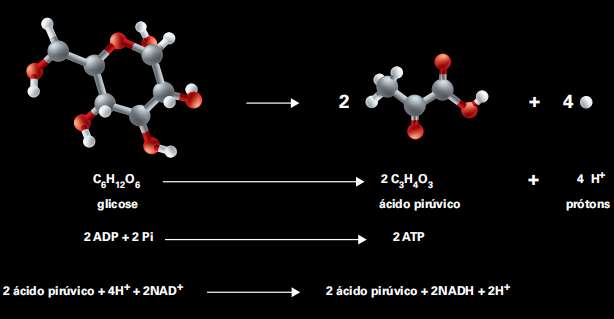
No processo denominado glicólise, uma molécula de glicose (C6H12O6 ) é convertida em duas moléculas de ácido pirúvico (C3H4O3 ), com liberação de energia, utilizada para a formação de 2 moléculas de ATP, e elétrons (formam-se 2 moléculas de NADH)
A fermentação láctica
O carregador de elétrons pode entregar o par de elétrons (e de prótons) ao próprio ácido pirúvico, que se transforma em ácido láctico. Nessa reação também há sobra de energia, que é utilizada para ligar ADP ao fosfato inorgânico, gerando moléculas de ATP.
Trata-se da fermentação láctica, via metabólica de baixa eficiência energética, pois cada molécula de glicose consumida leva à produção de apenas 2 moléculas de ATP. A fermentação não requer a participação das mitocôndrias e não consome oxigênio (por isso, é dita via anaeróbica), mas somente ácido pirúvico e NADH + H+, e não produz gás carbônico.
| NADH + H+ | + | C3H4O3 |
|
+ | C3H6O3 |
| NADH
carregado |
ácido pirúvico | NADH
descarregado |
ácido láctico |
Veja na figura abaixo uma representação esquemática de como as hemácias (ou glóbulos vermelhos do sangue) utilizam a energia da glicose depois que esta é convertida em ácido pirúvico. Embora não tenham nem mitocôndrias nem núcleo, no citoplasma dessas células estão presentes enzimas da via glicolítica (via de ocorrência da glicólise) e da fermentação láctica, que tornam possíveis esses fenômenos.
O ácido pirúvico é convertido a ácido láctico e não há produção de gás carbônico. Veja o esquema da reação:
NADH + H+ + CHO ➔ NAD+ + CHO
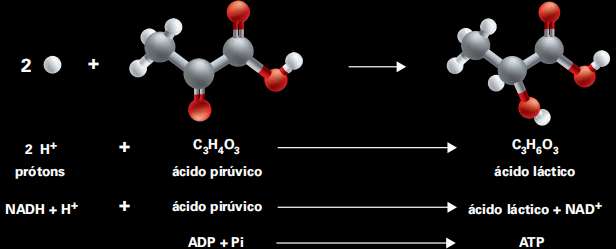
A fermentação láctica tem baixo rendimento energético, porém, garante a produção de ATP sem a necessidade de participação das mitocôndrias e sem consumo de oxigênio.
Observe que o carregador de elétrons participou da reação, sendo que dois elétrons e dois prótons foram deslocados para a molécula de ácido láctico que se formou.
Embora essa via metabólica alternativa seja pouco eficiente do ponto de vista energético, ela contribui para o aumento da eficiência das hemácias, pois lhes permite entregar a outras células do corpo humano todo o gás oxigênio que captaram em nossos pulmões (e que transportam ligado à hemoglobina), sem consumi-lo.
A fermentação alcoólica
Além da fermentação láctica, há outros tipos de fermentação, com a produção de substâncias diferentes. O fermento de padaria – um fungo unicelular na forma de levedura, também conhecido como levedo de cerveja –, por exemplo, realiza um tipo de fermentação em que há produção de álcool e gás carbônico.
A levedura de cerveja é um exemplo de organismo aeróbio não obrigatório, uma vez que, na presença de oxigênio, realiza respiração celular com suas mitocôndrias, mas, na ausência dessa substância (no interior da massa de pão, por exemplo), aciona a via da fermentação para a obtenção de energia e produz álcool e gás carbônico.
É possível reconhecer a produção de gás carbônico conforme as bolhas desse gás inflam a massa, que aumenta de volume. Quanto ao álcool, a maior parte evapora da massa enquanto ela descansa. Pergunte a um padeiro se ele não sente cheiro de álcool durante o preparo do pão!
Veja aqui no site uma aula completa sobre fermentação alcoólica.
Metabolismo energético II: mitocôndrias
O rendimento energético da glicose pode ser bem maior do que o obtido na fermentação láctica ou alcoólica. Para isso, no entanto, além dos insumos básicos
que se originam da glicólise, ou seja, o ácido pirúvico e os aceptores de elétrons, as células precisam também de um aceptor final para os elétrons, que venha de fora e seja capaz de ficar definitivamente com esses elétrons, levando-os embora. Esse aceptor final pode ser o gás oxigênio (respiração aeróbica), mas algumas bactérias e arqueas podem utilizar outras substâncias (respiração anaeróbica).
É necessária também uma infraestrutura física, que pode ser proporcionada por membranas celulares. Assim, algumas bactérias (células procarióticas) têm membranas dobradas do lado interno da parede bacteriana, e as células eucarióticas têm as mitocôndrias, organelas citoplasmáticas com dupla membrana, uma membrana externa e outra interna, esta bastante dobrada, o que aumenta muito sua superfície e os espaços inter membranas.
Acompanhe o que acontece em um organismo eucariótico. O ácido pirúvico entra na mitocôndria e participa de reações químicas que geram um novo substrato, com apenas dois átomos de carbono, um grupo acetil (C H O+).
Nessa reação também é produzido gás carbônico e há sobra de elétrons, que são recolhidos pelo aceptor de elétrons (o NAD+). Essa fase é chamada de oxidação do ácido pirúvico, que ocorre sem participação do oxigênio e resulta na produção de NADH + H+.
O grupo acetil – que possui dois átomos de carbono – gerado nessa etapa é muito reativo e logo se liga a uma molécula de coenzima A (C21H36N7O16P3S), substância abundante no interior da mitocôndria, formando o composto acetil-coenzima A.
Esse composto está pronto para participar de uma série de reações químicas cíclicas conhecidas como ciclo de Krebs – por terem sido elucidadas pelo bioquímico alemão Hans Krebs (1900-1981) – ou ciclo do ácido cítrico.
Essas reações, que não consomem oxigênio e não têm grande rendimento energético, produzem gás carbônico e coenzima-A, que pode voltar a participar do processo.
Mas a importância maior do ciclo de Krebs é o fato de, ao longo da série de reações que o constituem, sobrarem muitos elétrons, que são captados por aceptores de elétrons, como o NAD+ e o FAD. O ciclo de Krebs, portanto, é importante recarregador de aceptores de elétrons.
s reações, até aqui, ocorreram na parte central da mitocôndria (matriz mitocondrial). A partir deste ponto ocorrerão na superfície da membrana interna da organela, nas cristas mitocondriais.
Os aceptores de elétrons carregados se deslocam para a superfície dessa membrana, onde liberam os elétrons captados no ciclo de Krebs para proteínas de membrana, que têm o poder de receber o elétron e passá-lo adiante para outras proteínas, formando uma série ou cadeia.
Os elétrons passam de uma proteína à outra, em uma cadeia de quatro etapas. Quando o elétron perde um pouco de energia, ela é utilizada para fabricar ATP. Ao final, os elétrons já com baixa energia serão dirigidos ao oxigênio, daí essa etapa ser chamada cadeia respiratória, a única etapa em que há participação do oxigênio.
Como foi dito antes, em meio aquoso cada elétron consegue facilmente atrair um próton. A cada elétron liberado corresponde um próton, que é lançado no espaço inter membrana, represando nesse espaço as cargas positivas.
Em certos pontos da membrana, entretanto, os prótons podem retornar à matriz mitocondrial, e, ao fazê-lo, de maneira semelhante ao que acontece em um gerador de usina hidrelétrica, mas em proporções muito menores, eles literalmente giram um rotor molecular. Esse processo, que gera ATP e é denominado quimiosmose, está esquematizado na figura abaixo.
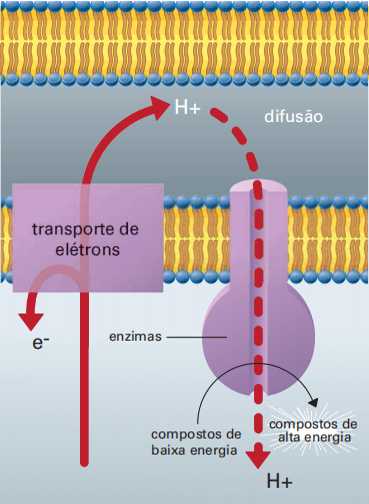
Os prótons se unem ao oxigênio que recebeu elétrons (reduzido) e assim é formada água.
A energia excedente é aproveitada para gerar moléculas de ATP, unindo difosfato de adenosina (ADP) ao fosfato inorgânico (Pi), na etapa denominada fosforilação oxidativa.
Assim, de uma única molécula de glicose, com a participação do oxigênio e graças à maquinaria do citoplasma e das mitocôndrias, são geradas 38 moléculas de ATP, a maior parte delas proveniente da cadeia respiratória e da quimiosmose. Veja um resumo do processo na tabela abaixo.
| RENDIMENTO ENERGÉTICO DE UMA MOLÉCULA DE GLICOSE | ||||
| Via metabólica | Glicólise | Oxidação do ácido pirúvico | Ciclo de Krebs | Cadeia respiratória
+ quimiosmose |
| Rendimento | 2 ATP | Nenhum | 2 ATP | 34 ATP |
| Produtos
energéticos |
2 NADH + H+ | NADH + H+ | NADH + H+
FADH2 |
Nenhum |
| Consumo de
oxigênio? |
Não | Não | Não | Não |
| Produtos de
descarte |
Nenhum | CO2 | CO2 | H2O |
Metabolismo energético III: respiração anaeróbica, quimiossíntese e fotossíntese
Respiração anaeróbica
Certos microrganismos, como bactérias e arqueas, vivem em condição anaeróbia, isto é, em meio pobre em oxigênio, mas, ainda assim, conseguem utilizar a energia de compostos alimentares de maneira mais eficiente do que a proporcionada pela fermentação.
Lembre-se de que as etapas em que ocorre maior produção de ATP na respiração celular são a cadeia respiratória e a quimiosmose.
Para isso, as células precisam de aceptores de elétrons carregados. Fala-se em respiração celular quando o destino final dos elétrons excedentes são compostos externos; se esse composto for o oxigênio, o processo é denominado respiração aeróbica, mas quando outras substâncias estão envolvidas, que não o oxigênio, falamos em respiração anaeróbica.
Existem arqueas e bactérias que vivem em ambientes sem oxigênio e conseguem aproveitar compostos de enxofre, nitrogênio, ferro ou, até mesmo, urânio como aceptores finais de elétrons; portanto, realizam respiração celular anaeróbica. Assim, a energia da matéria orgânica é aproveitada de maneira muito mais eficiente do que na fermentação, e também não há consumo de oxigênio.
Quimiossíntese
Existem microrganismos que, apesar de autótrofos, não dependem de luz para sobreviver. São procariotos que utilizam a energia de reações químicas para a síntese de carboidratos (matéria orgânica).
Daí o nome do processo, quimiossíntese. As arqueas metanogênicas vivem em ambiente com baixo teor de oxigênio e conseguem converter gás carbônico e hidrogênio em gás metano.
Elas estão presentes em pântanos, fundos oceânicos e também no sistema digestório de muitos animais, inclusive os seres humanos.
As bactérias do gênero Nitrobacter vivem no solo, em água doce e salgada, e em locais sem luz, e, na presença de oxigênio, oxidam compostos de nitrogênio, realizando uma quimiossíntese denominada nitrificação. Esse processo tem como resultado a formação de nitratos, que podem estar associados a diferentes metais (como sódio e potássio), mas a equação pode ser generalizada da seguinte forma:
NO – + 1/2 O ➔ NO –
Os nitritos são tóxicos para os seres vivos e essas bactérias desempenham um importante papel ecológico, em especial em locais poluídos, produzindo nitratos, que são menos tóxicos e facilmente utilizados por plantas e outros seres vivos.
Quando outras bactérias aeróbicas consomem oxigênio em locais poluídos, essa quimiossíntese cessa, devido à falta de um dos reagentes. Os níveis de oxigênio podem cair a ponto de haver um florescimento de bactérias anaeróbicas e a sobrevivência de animais, como peixes, por exemplo, se torna impossível. Isso explica mortandades repentinas em locais poluídos.
Fotossíntese
No passado, durante muito tempo, predominou a ideia de que as plantas retiravam seu sustento do solo. Como diferentes solos têm fertilidade diferente, pensava-se que as plantas absorvessem matéria do ambiente apenas pelas raízes.
Hoje em dia, entretanto, é consenso assumir que as plantas produzem seu próprio alimento, por meio da fotossíntese. A fotossíntese é um processo pelo qual as plantas, as algas e algumas bactérias produzem substâncias orgânicas a partir de substâncias inorgânicas do ambiente, como água (H2O) e gás carbônico (CO2), utilizando para isso a energia da luz.
A estrutura molecular de um pedaço de carvão vegetal revela muitos milhões de átomos de carbono que, antes de serem incorporados pela planta, estavam no ambiente. Por isso se diz que as plantas fixam carbono em seu organismo.
No entanto, por mais fértil que seja o solo de origem da planta, não é dele que elas retiram os átomos de carbono, como se acreditava no passado, e sim do ar atmosférico, ou seja, as plantas e outros organismos fotossintetizantes retiram gás carbônico da atmosfera.
No passado, há milhões de anos, as plantas que fixavam carbono do ambiente passaram por processos de fossilização. Combustíveis fósseis, como o carvão mineral e o petróleo, contêm o carbono retirado da atmosfera àquela época, período em que viveram os seres fotossintetizantes que deram origem a esses materiais.
Um pouco da história da fotossíntese
Em 1754 um cientista suíço chamado Charles Bonnet (1720-1793) realizou um experimento que se tornou clássico e que frequentemente é repetido em sala de aula.
Ao colocar em um local bem iluminado uma planta aquática (como as que são utilizadas em aquário), Bonnet observou que, após algum tempo, era possível perceber a formação de bolhas na água. Isso fez com que ele imaginasse que as plantas fabricavam algum “ar” – naquela época ainda não se sabia que o ar é uma mistura de gases.
Entre 1772-1773, o cientista e religioso inglês Joseph Priestley percebeu que as plantas tinham o poder de restaurar algumas qualidades do ar. Ele observara que a queima de uma vela em um ambiente fechado deixava esse ar impróprio para a respiração dos animais, e montou um experimento que comprovava a capacidade das plantas de restaurar as qualidades do ar, tornando-o novamente respirável.
Em 1782, Jean Senebier, um pastor suíço, comprovou que as plantas retiravam carbono do ar, mais especificamente do que na época era chamado “ar fixo” e que hoje denominamos gás carbônico. Além disso, ele estabeleceu que as plantas têm a capacidade de restaurar o poder comburente do ar. Ou seja, ficou comprovado que as
plantas produzem oxigênio, gás que alimenta a combustão e que só recebeu esse nome um pouco mais tarde, entre 1785-1786, batizado pelo químico francês Lavoisier.
No início do século XIX, os cloroplastos e a clorofila foram identificados como importantes para o metabolismo das plantas. E, já perto do final desse século, em 1883, por meio de um engenhoso experimento, Theodor W. Engelmann, com a ajuda de Carl Zeiss (1816-1888), meticuloso artesão de lentes, estabeleceu com precisão quais comprimentos de onda da luz são utilizados na fotossíntese.
Engelmann colocou uma alga verde filamentosa em água pobre em oxigênio e acrescentou ao meio bactérias aeróbicas. Em seguida, iluminou a alga com um feixe de luz branca que, ao atravessar um prisma, decompunha-se em diferentes comprimentos de onda, como em um arco-íris.
Observando a preparação ao microscópio, ele percebeu que as bactérias não se distribuíam igualmente ao longo do filamento da alga. Elas se concentravam nos segmentos da alga iluminados pelos comprimentos de onda relativos ao azul e ao vermelho.
Como as bactérias aeróbias dependem de oxigênio, elas se acumularam em áreas com maior concentração desse gás. Esses resultados mostraram que nem todos os comprimentos de onda são importantes para a fotossíntese, mas principalmente as frequências referentes ao azul e ao vermelho do espectro da luz visível.
No século XX a fotossíntese foi intensamente estudada. Algumas bactérias encontradas em lagos e fontes termais utilizam ácido sulfídrico em lugar de água para, na presença de luz solar, produzir carboidrato e enxofre.
O cientista C. B. van Niel (1897 – 1985) tinha demonstrado que o dióxido de carbono era reduzido por um composto oxidável – no caso, o ácido sulfídrico – e que essa era a origem do carboidrato nessas bactérias, de acordo com a seguinte equação resumida:
CO + 2 HS –> CH O + H O + 2 S
O esclarecimento desse processo, na década de 1930, indicava que deveria estar errada a suspeita da maioria dos cientistas à época, de que o gás oxigênio liberado na fotossíntese se originasse do gás carbônico.
Nos anos seguintes foram realizados experimentos cruciais para determinar se o oxigênio liberado na fotossíntese era proveniente do dióxido de carbono ou da água.
Em um primeiro experimento, pesquisadores colocaram algas em uma solução com água marcada com oxigênio radioativo e verificaram que o oxigênio produzido na fotossíntese apresentava radioatividade.
Pouco depois, em outro experimento, um grupo de cientistas forneceu gás carbônico marcado (radioativo) a algas e não foi encontrado traço de radioatividade no oxigênio produzido. Os experimentos esclareceram a pergunta inicial: o oxigênio produzido na fotossíntese provém da água, e não do dióxido de carbono.
Uma visão resumida
A fotossíntese pode ser entendida de maneira resumida como um conjunto de processos realizados por organismos autotróficos que podem utilizar a energia da luz para fabricar carboidratos a partir de gás carbônico e água, tendo como produto um carboidrato e como subprodutos o gás oxigênio e a água. A equação resumida da fotossíntese pode ser assim expressa:
| CO2 | + | 2 H2O | –> | CH2O | + O2 | + | H2O |
| gás carbônico | + | água | –> | carboidrato | + gás oxigênio | + | água |
Essa equação resumida, amplamente conhecida, levou muitas pessoas a considerar que a fotossíntese seria simplesmente o processo inverso da respiração celular.
Outra confusão comum a respeito é acreditar que os dois processos são mutuamente excludentes, ou seja, que a planta que realiza fotossíntese não poderia realizar respiração celular. Nenhuma dessas suposições tem fundamento.
A respiração celular ocorre continuamente nos organismos que realizam fotossíntese, seja de dia, seja à noite. A fotossíntese depende da presença de luz, mas, enquanto ela é realizada, a respiração celular não cessa.
A fotossíntese é muito diferente da respiração celular, pois produz moléculas de carboidratos que serão utilizadas por todas as células que realizam respiração aeróbica, tanto animais quanto vegetais.
Durante o dia, em especial na presença de muita luz, a fotossíntese produz mais carboidratos e oxigênio do que o próprio organismo consome na respiração. Daí haver excedentes, que são armazenados na forma de amido, no caso dos carboidratos, ou simplesmente eliminados para o ambiente, no caso do oxigênio.
A fotossíntese pode ser realizada por organismos procariotos e eucariotos, mas só estes últimos têm organelas especializadas para esse fim, os cloroplastos.
Como você estudou no capítulo anterior, as biomembranas têm propriedades que lhes permitem armazenar e transferir energia. A exemplo das mitocôndrias, os cloroplastos são organelas com membranas que delimitam um complexo sistema de compartimentos, (os tilacoides) nos
quais estão os pigmentos sensíveis à luz, como a clorofila. Os cloroplastos têm membrana dupla, e, em seu interior, no estroma, há DNA, bem como enzimas para síntese de aminoácidos e proteínas.
Os tilacoides formam pilhas (grana), e em seu interior (lúmen do tilacoide) ocorrem diversas reações químicas, como veremos a seguir.
Os cloroplastos são componentes típicos das células das plantas, mas ocorrem também em algas e em muitos seres unicelulares. As bactérias que realizam fotossíntese não possuem cloroplastos.
Diversas descobertas ao longo do século XX permitiram compreender que a fotossíntese é um conjunto de processos que ocorrem em duas etapas básicas distintas. Inicialmente a luz é captada pela clorofila, presente nas membranas dos tilacoides, e parte dessa energia é utilizada para transferir elétrons a outras substâncias (aceptores de elétrons).
A clorofila recupera seus elétrons com muita avidez, a partir da oxidação da água com a ajuda de compostos que possuem manganês, que acumulam cargas positivas. Quando esses compostos perdem elétrons, sua carga positiva aumenta a ponto de retirar elétrons da molécula da água.
Ela se torna instável, até se desagregar em um processo denominado fotólise. Dessa quebra resultam elétrons de alta energia, que são capturados por compostos da membrana, e prótons (H+), que se acumulam no lúmen do tilacoide. Além de prótons e elétrons, é liberado oxigênio molecular (O2), que deixa o cloroplasto.
Os elétrons são carreados por substâncias imersas na membrana e chegam a um complexo onde também há clorofila. A ação da luz solar levará a incorporar os elétrons a uma substância muito semelhante à que ocorre nas mitocôndrias, o NAD+, mas que possui um átomo de fósforo, daí sua abreviatura NADP+.
Esse carregador de elétrons leva consigo, além do elétron, um próton, como vimos no caso da respiração celular. Nessa região da membrana, o NADP+ se transforma em NADPH.
O acúmulo de prótons no interior do tilacoide o torna bastante ácido, em contraste com o estroma do cloroplasto. Essa abundância de prótons é aproveitada para regenerar ATP, em um processo de adição de um grupo fosfato (fosforilação) muito semelhante ao que ocorre na cadeia respiratória das mitocôndrias, a quimiosmose.
Ao passar pela membrana do tilacoide os prótons literalmente giram um rotor molecular, que permite acoplar fosfato inorgânico a ADP, o que gera ATP (reveja o item 5, Metabolismo energético II: mitocôndrias).
Uma parte desse ATP será utilizada na etapa seguinte da fotossíntese e em diversos outros processos que ocorrem no interior dos cloroplastos, como a síntese de aminoácidos a partir de carboidratos.
Na segunda etapa do processo, a energia do ATP produzido, os elétrons e os prótons (H+) que os acompanham no aceptor reduzido (NADPH) serão utilizados em uma sequência de reações químicas, que reduzem o dióxido de carbono (CO2), tendo como resultado carboidrato (CH2O).
Um conjunto de reações químicas, conhecido como ciclo de Calvin (ou ciclo das pentoses), ocorre no estroma do cloroplasto – ou seja, fora do tilacoide –, demanda dióxido de carbono e resulta em carboidrato. Essas reações independem da luz e, por isso, constituem a chamada fase escura da fotossíntese, que consome dióxido de carbono e produz carboidrato.
No ciclo de Calvin, além do carboidrato, são gerados compostos de baixa energia (ADP + Pi e NADP+), que são recarregados nas reações que dependem de luz (ou fase clara da fotossíntese). Embora tradicionalmente a fotossíntese seja apresentada como um processo que tem a glicose como produto final, é preciso lembrar que isso decorre apenas de uma simplificação didática do processo.
Na realidade, pouca glicose livre é produzida pelas células fotossintetizantes. A maior parte do carbono fixado é convertida em sacarose, a principal forma de carboidrato transportada nos vegetais (lembre-se de sua grande solubilidade em água), ou em amido, que não requer água para ser armazenado (é pouco solúvel em água).
A fotossíntese é realizada por diversos organismos, eucariotos unicelulares e pluricelulares, e também por diversos procariotos. No entanto, nem animais nem fungos são capazes de realizar a fotossíntese.